Примером анаэробного дыхания является. Анаэробное дыхание
(в редких случаях - и эукариотами) в анаэробных условиях. При этом факультативные анаэробы используют акцепторы электронов с высоким окислительно-восстановительным потенциалом (NO 3 − , NO 2 − , Fe 3+ , фумарат , диметилсульфоксид и т. д.), у них это дыхание конкурирует с энергетически более выгодным аэробным и подавляется кислородом. Акцепторы с низким окислительно-восстановительным потенциалом (сера , SO 4 2− , CO 2) применяются только строгими анаэробами, гибнущими при появлении в среде кислорода. В корневых системах многих растений при гипоксии и аноксии, вызванных затоплением посевов в результате длительных дождей или весенних паводков, развивается анаэробное дыхание с использованием в качестве акцепторов электронов альтернативных кислороду соединений, например нитратов. Установлено, что растения, произрастающие на полях, удобренных нитратными соединениями, переносят переувлажнение почвы и сопутствующую ему гипоксию лучше, нежели такие же растения без нитратной подкормки.
Механизмы окисления органических субстратов при анаэробном дыхании, как правило, аналогичны механизмам окисления при аэробном дыхании. Исключением является использование в качестве исходного субстрата ароматических соединений. Обычные пути их катаболизма требуют молекулярного кислорода уже на первых стадиях, в анаэробных условиях осуществляются иные процессы, например, восстановительная деароматизация бензоил-КоА у Thauera aromatica с затратой энергии АТФ. Некоторые субстраты (например, лигнин) при анаэробном дыхании не могут использоваться.
Нитратное и нитритное дыхание
Больше путей переноса протонов через мембрану анаэробная ЭТЦ не содержит (в аэробной же их 3), в связи с чем нитратное дыхание по эффективности в расчёте на 1 моль глюкозы составляет лишь 70 % от аэробного. При поступлении в среду молекулярного кислорода бактерии переключаются на обычное дыхание.
Нитратное дыхание встречается, хотя и редко, и среди эукариот. Так, нитратное дыхание, сопровождающееся денитрификацией и выделением молекулярного азота, недавно открыто у фораминифер . До этого нитратное дыхание с образованием N 2 O было описано у грибов Fusarium и Cylindrocarpon (см. .
Сульфатное дыхание
В настоящее время известен ряд бактерий, способных окислять органические соединения или молекулярный водород в анаэробных условиях, используя в качестве акцепторов электронов в дыхательной цепи сульфаты , тиосульфаты , сульфиты , молекулярную серу . Этот процесс получил название диссимиляционной сульфатредукции, а бактерии, осуществляющие этот процесс - сульфатвосстанавливающих или сульфатредуцирующих.
Все сульфатвосстанавливающие бактерии - облигатные анаэробы.
Сульфатвосстанавливающие бактерии получают энергию в процессе сульфатного дыхания при переносе электронов в электронтранспортной цепи. Перенос электронов от окисляемого субстрата по электронтранспортной цепи сопровождается возникновением электрохимического градиента ионов водорода с последующим синтезом АТФ .
Подавляющее большинство бактерий этой группы хемоорганогетеротрофы. Источником углерода и донором электронов для них являются простые органические вещества - пируват , лактат , сукцинат , малат , а также некоторые спирты . У некоторых сульфатвосстанавливающих бактерий обнаружена способность к хемолитоавтотрофии, когда окисляемым субстратом является молекулярный водород .
Сульфатвосстанавливающие эубактерии широко распространены в анаэробных зонах водоёмов разного типа, в иле , в почвах , в пищеварительном тракте животных . Наиболее интенсивно восстановление сульфатов происходит в соленых озерах и морских лиманах , где почти нет циркуляции воды, и содержится много сульфатов. Сульфатвосстанавливающим эубактериям принадлежит ведущая роль в образовании сероводорода в природе и в отложении сульфидных минералов . Накопление в среде H 2 S часто приводит к отрицательным последствиям - в водоемах к гибели рыбы , в почвах к угнетению растений . С активностью сульфатвосстанавливающих эубактерий связана также коррозия в анаэробных условиях различного металлического оборудования, например, металлических труб.
Фумаратное дыхание
В качестве акцептора электронов может использоваться фумарат . Фумаратредуктаза сходна с нитритредуктазой: лишь вместо молибдоптерин содержащей субъединицы в её состав входит ФАД и гистидин содержащая субъединица. Трансмембранный протонный потенциал образуется аналогичным образом: перенос протонов не происходит, однако фумаратредуктаза связывает протоны в цитоплазме, а дегидрогеназы в начале ЭТЦ выделяют протоны в периплазму. Перенос электронов с дегидрогеназ на фумаратредуктазу происходит обычно через мембранный пул менохинонов.
Фумарат, как правило, отсутствует в природных местообитаниях и образуется самими микроорганизмами из аспартата , аспарагина , сахаров , малата и цитрата . В виду этого большинство бактерий, способных к фумаратному дыханию содержат фумаразу, аспартат:аммиак-лиазу и аспарагиназу , синтез которого контролирует чувствительный к молекулярному кислороду белок Fnr.
Фумаратное дыхание достаточно широко распространено среди эукариот, в частности у животных (среди животных, у которых оно описано - пескожил , мидии , аскарида , печеночная двуустка и др .)
Железистое дыхание
Дыхание ацетогенных бактерий
Строго анаэробные ацетогенные бактерии родов Acetobacterium , Clostridium , Peptostreptococcus и др. способны получать энергию, окисляя водород углекислым газом. При этом две молекулы CO 2 образуют ацетат . Энергия при этом запасается в виде трансмембранного градиента протонов (Clostridium sp. ) или ионов натрия (Acetobacterium woodi ). Для перевода его в энергию связей АТФ используется обычная H-транспортирующая АТФ-синтаза или Na-зависимая АТФ-синтаза соответственно.
Анаэробное дыхание у растений
Анаэробное дыхание , в частности нитратное , активируется в корневых системах некоторых растений в условиях аноксии и гипоксии . Однако если у многих бактерий и некоторых протистов и животных оно может быть основным и достаточным для получения энергии процессом (часто наряду с гликолизом), то у растений функционирует почти исключительно в стрессовых условиях. Так или иначе, но на полях, где в качестве удобрений были внесены нитраты , растения лучше переносят гипоксию , вызванную переувлажнением почвы из-за продолжительных дождей .
Анаэробное дыхание у грибов, протистов и животных
Среди животных анаэробное фумаратное дыхание встречается у некоторых паразитических и свободноживущих червей, ракообразных, моллюсков; нитратное дыхание известно среди грибов (например, у Fusarium)
Аэробное дыхание (терминальное окисление, или окислительное фосфорилирование) – это совокупность катаболитических процессов на мембранах митохондрий, завершающихся полным окислением органических веществ с участием молекулярного кислорода. При этом роль протонного резервуара играет межмембранный матрикс – пространство между внешней и внутренней мембранами.
Электроны, потерявшие энергию, поступают на комплекс ферментов под названием цитохромоксидаза. Цитохромоксидаза использует электроны для активации (восстановления) молекулярного кислорода О 2 до О 2 2– . Ионы О 2 2– присоединяют протоны, образуя пероксид водорода, который при помощи каталазы разлагается на Н 2 О и О 2 . Последовательность описанных реакций можно представить в виде схемы:
2О 2 + 2ē → 2О 2 2– ; 2О 2 2– + 4Н + → 2Н 2 О 2 ; 2Н 2 О 2 → 2Н 2 О + О 2
Суммарное уравнение аэробного дыхания:
С 6 Н 12 О 6 + 6 О 2 + 38 АДФ + 38 Ф → 6 СО 2 + 6 Н 2 О + 38 АТФ + Q
Вопросы для самоконтроля
1.В чем сущность процесса дыхания?
2.Каково суммарное уравнение процесса дыхания?
3.В чем состоит окислительное фосфорилирование?
4.В чем заключается гликолиз?
5.Что охватывает цикл Кребса?
6.Чем характеризуются анаэробное дыхание и спиртовое брожение?
7.Как происходят масляно-кислое и молочно-кислое брожения? Где они встречаются?
8. Какова энергетическая сторона процесса дыхания и процесса брожения?
9. Какие опыты доказывают наличие процесса дыхания у растений?
10. Что называется дыхательным коэффициентом?
ЛЕКЦИЯ 6
Тема: Потребность растений в элементах минерального питания. Макроэлементы, микроэлементы. Питательные смеси для культивирования растений и изолированных клеток. Взаимодействие ионов. Особенности почвы как питающего растения субстрата. Проникновение ионов в растительную клетку. Активный и пассивный транспорт ионов через мембрану.
Цель лекции: Показать потребность растений в элементах минерального питания. Питательные смеси для культивирования растений и изолированных клеток, макроэлементы, микроэлементы. Активный и пассивный транспорт ионов через мембрану.
Минеральное питание – это поглощение минеральных веществ в виде ионов, их передвижение по растению и включение в обмен веществ . В составе растений обнаружены почти все существующие на Земле химические элементы. Элементы питания поглощаются из воздуха - в форме углекислого газа (CO 2) и из почвы - в форме воды (H 2 O) и ионов минеральных солей. У высших наземных растений различают воздушное, или листовое, питание (Фотосинтез ) и почвенное, или корневое, питание (Минеральное питание растений ). Низшие растения (бактерии, грибы, водоросли) поглощают CO 2 , H 2 O и соли всей поверхностью тела.
Почва является необходимым и незаменимым субстратом, в котором растения укрепляются своими корнями, и из которого черпают влагу и элементы минерального питания. Велика роль почвы в формировании и сохранении биологического разнообразия.
С другой стороны - потоки всех элементов в биосфере проходят через почву, которая посредством специфических механизмов регулирует их направленность и интенсивность.
Одноклеточные организмы и водные растения поглощают ионы всей поверхностью, высшие наземные растения - поверхностными клетками корня , в основном корневыми волосками .
Через корень растения поглощают из почвы главным образом ионы минеральных солей, а также некоторые продукты жизнедеятельности почвенных микроорганизмов и корневые выделения др. растений. Ионы сначала адсорбируются на клеточных оболочках, затем проникают в цитоплазму через плазмалемму. Катионы (за исключением К+) проникают через мембрану пассивно, путём диффузии, анионы, а также К + (при низких концентрациях) - активно, с помощью молекулярных «ионных насосов», транспортирующих ионы с затратой энергии. Каждый элемент минерального питания играет в обмене веществ определённую роль и не может быть полностью заменен др. элементом. Анализ сухого вещества растений показывает, что в нем содержатся углерод (45 %), кислород (42 %), водород (6,5 %), азот (1,5 %), зольные элементы (5 %).
Все элементы, встречающиеся в растениях, принято делить на три группы:
Макроэлементы . 2. Микроэлементы. 3. Ультрамикроэлементы .
Ионы, поступающие в раст. клетку вступают в определенные взаимодействия, и типы этих взаимодействий различны.
Различают такие типы взаимодействий как антогонизм, синергизм, аддитивность.
Антогонизм ионов – это снижение одними катионами ядовитого эффекта других, обусловленного их взаимодействием с коллоидами протоплазмы. Синергизм - это комбинированное воздействие двух или более ионов, характеризующееся тем, что их объединённое биологическое действие существенно превосходит эффект каждого отдельно взятого компонента. Аддитивность – эффект совместного действия ионов равный сумме эффектов действия каждого вещества в отдельности.
В естественных условиях, растения получают необходимые вещества непосредственно из почвы, через корневую систему. В искусственных условиях чаще всего для выращивания растений используют метод гидропоники. Гидропоника (от гидро… и греч. pónos - работа) - выращивание растений не в почве, а в специальном питательном растворе. Питательный раствор представляет собой водный раствор веществ, необходимых растению для жизни и роста. При гидропонном методе выращивания растений все элементы должны содержаться в питательном растворе в оптимальном количестве.
Вопросы для самоконтроля
1. Какие элементы являются органогенами, их процентное содержание в сухом веществе растения?
2. Какие зольные микроэлементы вы знаете? Какова их роль в растении?
3. Какие микроэлементы вам известны? Какую роль они играют в жизни растений?
4. В чем сущность нитрификации и денитрификации?
5. Дайте общую характеристику макро и микроэлементов.
6. Типы взаимодействия ионов в растительных клетках: синергизм, аддитивность, антогогизм.
1. Определение понятия «анаэробное дыхание».
2. Нитратное дыхание.
3. Сульфатное дыхание.
4. Карбонатное дыхание.
5. Фумаратное дыхание.
1. Определение понятия «анаэробное дыхание»
Анаэробное дыхание – цепь анаэробных окислительно- восстановительных реакций, которые сводятся к окислению органического или неорганического субстрата с использованием в качестве конечного акцептора электронов не молекулярного кислорода, а других неорганических веществ (нитрата –NO 3 - , нитрита – NO 2 - , сульфата – SO 4 2- , сульфита – SO 3 2- , CO 2 и др.), а также органических веществ (фумарата и др.). Молекулы АТФ в процессе анаэробного дыхания образуются в основном в электронтранспортной цепи, т. е. в результате реакций мембранного фосфорилирования, но в меньшем количестве, чем при аэробном дыхании.
При анаэробном дыхании конечным акцептором электронов в электронтранспортной цепи являются неорганические или органические соединения. Например, если конечным акцептором электронов является SO 4 2- , то процесс называют сульфатным дыханием , а бактерии – с ульфатвосстанавливающими или сульфатредуцирующими . В том случае, если конечным акцептором электронов служит NO 3 - или NO 2 - , то процесс называется нитратным дыханием или денитрификацией , а бактерии, осуществляющие этот процесс, – денитрифицирующими . В качестве конечного акцептора электронов может выступать СО 2. . Процесс соответственно называют карбонатным дыханием , а бактерии – метаногенными (метанобразующими ) . Одним из немногих примеров, когда конечным акцептором служит органическое вещество, является фумаратное дыхание .
Основные особенности бактерий, способных к анаэробному дыханию :
1) Имеют укороченные электронтранспортные, или дыхательные, цепи, т. е. они не содержат всех переносчиков, характерных для дыхательных цепей, функционирующих в аэробных условиях.
2) В дыхательных цепях анаэробов цитохромоксидаза заменена соответствующими редуктазами.
3) У строгих анаэробов не функционирует цикл Кребса или же он разорван и выполняет только биосинтетические функции.
4) Основное количество молекул АТФ при анаэробном дыхании синтезируется в процессе мембранного фосфорилирования.
5) По отношению к молекулярному кислороду бактерии, осуществляющие анаэробное дыхание, являются факультативными или облигатными анаэробами. К облигатным анаэробам относятся сульфатвосстанавливающие и метаногенные бактерии. К факультативным анаэробам – денитрифицирующие бактерии и бактерии, осуществляющие фумаратное дыхание. Факультативные анаэробы могут переключать свой энергетический метаболизм с аэробного дыхания в присутствии в среде молекулярного кислорода на анаэробное дыхание в отсутствии молекулярного кислорода.
6) Выход АТФ при анаэробном дыхании меньше, чем при аэробном, но больше, чем при брожении.
2. Нитратное дыхание
Конечными акцепторами электронов при нитратном дыхании являются нитраты (NO 3 - ) или нитриты (NO 2 -). Результатом нитратного дыхания является восстановление NO 3 - или NO 2 - до газообразных продуктов (NО, N 2 O, N2). Суммарную реакцию нитратного дыхания , где окисляемым субстратом является глюкоза, а конечным акцептором электронов – нитраты, можно записать следующим образом:
С 6 Н 12 О 6 + 4NО - 3 → 6СО 2 + 6Н 2 О + 2N 2 + х(кДж)
Полный процесс денитрификации состоит из четырех реакций восстановления, каждая из которых катализируется специфическими мембраносвязанными редуктазами.
Первый этап : восстановление нитрата до нитрита, катализируют молибденсодержащие ферменты нитратредуктазы:
NО - 3 + 2е - + 2Н + → NО - 2 + Н 2 О
Второй этап : восстановление нитрита до оксида азота, катализируют нитритредуктазы:
NО - 2 + е - + Н + → NО + ОН -
Нитрат- и нитритредуктазы очень чувствительны к молекулярному кислороду, который ингибирует их активность, а также репрессирует синтез. Соответственно данные реакции могут протекать только в том случае, когда кислород полностью отсутствует или когда его концентрация незначительна.
Третий этап : восстановление оксида азота до закиси азота, катализируют редуктазы оксида азота:
2NО + 2е - + 2Н + → N 2 О + Н 2 О
Четвертый этап : восстановление закиси азота в молекулярный азот, катализируют редуктазы закиси азота:
N 2 О + 2е - + 2Н + → N 2 + Н 2 О
У денитрифицирующих бактерий, которые являются факультативными анаэробами, функционирует полная электронтранспортная цепь в случае аэробного дыхания, и укороченная – при анаэробном дыхании.
Электронтранспортные цепи бактерий-денитрификаторов в анаэробных условиях содержат все основные типы связанных с мембранами переносчиков: флавопротеины, хиноны, цитохромы b и с . Установлено, что нитратредуктазы денитрифицирующих бактерий связаны с дыхательной цепью на уровне цитохрома b , а нитритредуктазы и редуктазы оксида азота и закиси азота на уровне цитохрома с. Процесс полной денитрификации, когда происходит восстановление NO 3 - до N 2, транспорт электронов в дыхательной цепи можно представить следующим образом (рис. 1).
Рис. 1. Транспорт электронов в процессе денитрификации
Количество синтезируемых молекул АТФ зависит от строения дыхательной цепи, наличия и свойств соответствующих редуктаз. При «полной» денитрификации энергии запасается в большем количестве, чем при «усеченной», когда осуществляются только отдельные этапы этого процесса:
NО - 3 → NО - 2 ; NО → N 2 О; N 2 О → N 2 .
Схематично нитратное дыхание при окислении глюкозы можно представить следующим образом (рис. 2).
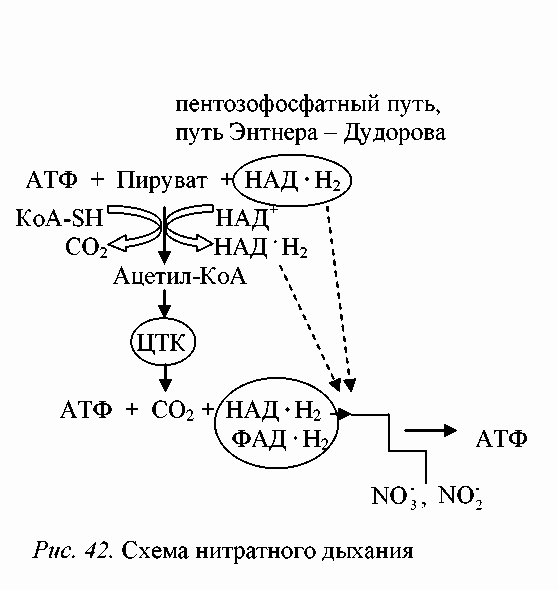
Рис. 2. Схема нитратного дыхания
Распространение и роль денитрифицирующих бактерий. Денитрифицирующие бактерии широко распространены в природе. Они принадлежат ко всем основным физиологическим группам: фототрофным, хемолитотрофным, грамположительным и грамотрицательным факультативным анаэробам. Однако в большей степени способность к денитрификации распространена у бактерий родов Bacillus и Pseudomonas .
Денитрифицирующие бактерии – это обитатели пресных и морских водоемов, почв разного типа, хотя процесс денитрификации у них происходит только в анаэробных условиях. Процесс денитрификации считается вредным для сельского хозяйства, так как доступные для растений нитраты превращаются в недоступный для них молекулярный азот, что приводит к обеднению почвы азотом. Тем не менее, денитрифицирующие бактерии являются важным звеном в круговороте азота в природе, обогащающим атмосферу молекулярным азотом. Кроме того, эти бактерии играют положительную роль в очистке подземных вод и почв от накопившихся в результате деятельности человека (внесение высоких доз удобрений, промышленные стоки) нитратов и нитритов, которые в больших концентрациях токсичны для живых организмов. В связи с этим денитрифицирующие бактерии используют для очистки сточных вод от нитратов.
Введение
1. Аэробное дыхание
1.1 Окислительное фосфолирование
2. Анаэробное дыхание
2.1 Типы анаэробного дыхания
4.Список литературы
Введение
Дыхание присуще всем живым организмам. Оно представляет собой окислительный распад органических веществ, синтезированных в процессе фотосинтеза, протекающих с потреблением кислорода и выделением диоксида углерода. А.С. Фаминцын рассматривал фотосинтез и дыхание как две последовательные фазы питания растений: фотосинтез готовит углеводы, дыхание перерабатывает их в структурную биомассу растения, образуя в процессе ступенчатого окисления реакционноспособные вещества и освобождая энергию, необходимую для их превращения и процессов жизнедеятельности в целом. Суммарное уравнение дыхания имеет вид:
CHO + 6O → 6CO + 6HO + 2875кДж.
Из этого уравнения становится ясно, почему именно скорость газообмена используют для оценки интенсивности дыхания. Оно было предложено в 1912 г. В. И. Палладиным, который считал, что дыхание состоит из двух фаз - анаэробной и аэробной. На анаэробном этапе дыхания, идущем в отсутствие кислорода, глюкоза окисляется за счет отнятия водорода (дегидрирования), который, по мнению ученого, передается на дыхательный фермент. Последний при этом восстанавливается. На аэробном этапе происходит регенерация дыхательного фермента в окислительную форму. В. И. Палладин впервые показал, что окисление сахара идет за счет непосредственного окисления его кислородом воздуха, поскольку кислород не встречается с углеродом дыхательного субстрата, а связано с его дегидрированием.
Существенный вклад в изучение сути окислительных процессов и химизма процесса дыхания внесли как отечественные (И.П. Бородин, А.Н.Бах, С.П. Костычев, В.И. Палладин), так и зарубежные (А.Л. Лавуазье, Г. Виланд, Г. Кребс) исследователи.
Жизнь любого организма неразрывно связана с непрерывным использованием свободной энергии, генерируемой при дыхании. Неудивительно, что изучению роли дыхания в жизни растения в последнее время отводят центральное место в физиологии растений.
1. Аэробное дыхание
Аэробное дыхание - это окислительный процесс, в ходе которого расходуется кислород. При дыхании субстрат без остатка расщепляется до бедных энергией неорганических веществ с высоким выходом энергии. Важнейшими субстратами для дыхания служат углеводы. Кроме того, при дыхании могут расходоваться жиры и белки.
Аэробное дыхание включает два основных этапа:
- бескислородный, в процессе, которого происходит постепенное расщепление субстрата с высвобождением атомов водорода и связыванием с коферментами (переносчиками типа НАД и ФАД);
- кислородный, в ходе которого происходит дальнейшее отщепление атомов водорода от производных дыхательного субстрата и постепенное окисление атомов водорода в результате переноса их электронов на кислород.
На первом этапе вначале высокомолекулярные органические вещества (полисахариды, липиды, белки, нуклеиновые кислоты и др.) под действием ферментов расщепляются на более простые соединения (глюкозу, высшие карбоновые кислоты, глицерол, аминокислоты, нуклеотиды и т.п.) Этот процесс происходит в цитоплазме клеток и сопровождается выделением небольшого количества энергии, которая рассеивается в виде тепла. Далее происходит ферментативное расщепление простых органических соединений.
Примером такого процесса является гликолиз - многоступенчатое бескислородное расщепление глюкозы. В реакциях гликолиза шестиуглеродная молекула глюкозы (С) расщепляется на две трехуглеродные молекулы пировиноградной кислоты (С). При этом образуется две молекулы АТФ, и выделяются атомы водорода. Последние присоединяются к переносчику НАД (никотинамидадениндинклеотид), который переходит в свою восстановительную форму НАД ∙ Н + Н. НАД кофермент, близкий по своей структуре к НАДФ. Оба они представляют собой производные никотиновой кислоты - одного из витаминов группы В. Молекулы обоих коферментов электроположительны (у них отсутствует один электрон) и могут играть роль переносчика как электронов, так и атомов водорода. Когда акцептируется пара атомов водорода, один из атомов диссоциирует на протон и электрон:
а второй присоединяется к НАД или НАДФ целиком:
НАД+ Н + [Н+ е] → НАД ∙ Н + Н.
Свободный протон позднее используется для обратного окисления кофермента. Суммарно реакция гликолиза имеет вид
CHO +2АДФ + 2НРО + 2 НАД→
2СНО + 2АТФ + 2 НАД ∙ Н + Н+ 2 HO
Продукт гликолиза - пировиноградная кислота (СНО) - заключает в себе значительную часть энергии, и дальнейшее ее высвобождение осуществляется в митохондриях. Здесь происходит полное окисление пировиноградной кислоты до COи HO. Этот процесс можно разделить на три основные стадии:
- окислительное декарбоксилирование пировиноградной кислоты;
- цикл трикарбоновых кислот (цикл Кребса);
- заключительная стадия окисления - электронтранспортная цепь.
На первой стадии пировиноградная кислота взаимодействует с веществом, которое называют коферментом А, в результате чего образуется ацетилкофермент а с высокоэнергетической связью. При этом от молекулы пировиноградной кислоты отщепляется молекула CO(первая) и атомы водорода, которые запасаются в форме НАД ∙ Н + Н.
Вторая стадия - цикл Кребса (рис. 1)
В цикл Кребса вступает ацетил-КоА, образованный на предыдущей стадии. Ацетил-КоА взаимодействует со щавелево-уксусной кислотой, в результате образуется шестиуглеродная лимонная кислота. Для этой реакции требуется энергия; ее поставляет высокоэнергетическая связь ацетил-КоА. В конце цикла щавелево-лимонная кислота регенерируется в прежнем виде. Теперь она способна вступить в реакцию с новой молекулой ацетил-КоА, и цикл повторяется. Суммарно реакция цикла может быть выражена следующим уравнением:
ацетил-КоА + 3HO + 3НАД+ ФАД + АДФ + НРО→
КоА + 2CO+ 3НАД ∙ Н + Н+ФАД ∙ H+ АТФ.
Таким образом, в результате распада одной молекулы пировиноградной кислоты в аэробной фазе (декарбоксилирование ПВК и цикла Кребса) выделяется 3CO, 4 НАД ∙ Н + Н, ФАД ∙ H. Суммарно реакцию гликолиза, окислительного декарбоксилирования и цикла Кребса можно записать в следующем виде:
CHO + 6 HO + 10 НАД + 2ФАД →
6CO+ 4АТФ + 10 НАД ∙ Н + Н+ 2ФАД ∙ H.
Третья стадия - электротранспортная цепь.
Пары водородных атомов, отщепляемые от промежуточных продуктов в реакциях дегидрирования при гликолизе и в цикле Кребса, в конце концов, окисляются молекулярным кислородом до HO с одновременным фосфолированием АДФ в АТФ. Происходит это тогда, когда водород, отделившийся от НАД ∙ Hи ФАД ∙ H, передается по цепи переносчиков, встроенных во внутреннюю мембрану митохондрий. Пары атомов водорода 2Н можно рассматривать как 2 Н + 2е. Движущей силой транспорта атомов водорода в дыхательной цепи является разность потенциалов.
С помощью переносчиков ионы водорода Нпереносятся с внутренней стороны мембраны на ее внешнюю сторону, иначе говоря, из матрикса митохондрии в межмембранное пространство (рис. 2).
|
|
При переносе пары электронов от над на кислород они пересекают мембрану три раза, и этот процесс сопровождается выделением на внешнюю сторону мембраны шести протонов. На заключительном этапе протоны переносятся на внутреннюю сторону мембраны и акцептируются кислородом:
½ O + 2е → O.
В результате такого переноса ионов Нна внешнюю сторону мембраны митохондрий в перимитохондриальном пространстве создается концентрация их, т.е. возникает электрохимический градиент протонов.
Когда протонный градиент достигает определенной величины, ионы водорода из Н-резервуара движутся по специальным каналам в мембране, и их запас энергии используется для синтеза АТФ. В матриксе они соединяются с заряженными частичками О, и образуется вода: 2Н+ О²ˉ → HO.
1.1 Окислительное фосфолирование
Процесс образования АТФ в результате переноса ионов Нчерез мембрану митохондрии получил название окислительного фосфолирования. Он осуществляется при участии фермента АТФ-синтетазы. Молекулы АТФ-синтетазы располагаются в виде сферических гранул на внутренней стороне внутренней мембраны митохондрий.
В результате расщепления двух молекул пировиноградной кислоты и переноса ионов водорода через мембрану по специальным каналам синтезируется в целом 36 молекул АТФ (2 молекулы в цикле Кребса и 34 молекулы в результате переноса ионов Н через мембрану).
Суммарное уравнение аэробного дыхания можно выразить следующим образом:
CHO + O+ 6HO + 38АДФ + 38НРО→
6CO+ 12HO + 38АТФ
Совершенно очевидно, что аэробное дыхание прекратится в отсутствии кислорода, поскольку именно кислород служит конечным акцептором водорода. Если клетки не получают достаточного количества кислорода, все переносчики водорода вскоре полностью насытятся и не смогут передавать его дальше. В результате основной источник энергии дл образования АТФ окажется блокированным.
аэробное дыхание окисление фотосинтез
2. Анаэробное дыхание
Анаэробное дыхание. Некоторые микроорганизмы способны использовать для окисления органических или неорганических веществ не молекулярный кислород, а другие окисленные соединения, например, соли азотной, серной и угольной кислот, превращающиеся при этом в более восстановленные соединения. Процессы идут в анаэробных условиях, и их называют анаэробным дыханием:
2HNO + 12Н→ N + 6HO + 2Н
HSO + 8Н→ HS + 4HO
У микроорганизмов, осуществляющих такое дыхание, конечным акцептором электронов будет не кислород а неорганическое соединения - нитриты, сульфаты и карбонаты. Таким образом, различия между аэробным и анаэробным дыханием заключается в природе конечного акцептора электронов.
2.1 Типы анаэробного дыхания
Основные типы анаэробного дыхания приведены в таблице 1. есть также данные об использовании бактериями в качестве акцепторов электронов Mn, хроматов, хинонов и др.
Таблица 1 Типы анаэробного дыхания у прокариот (по: М.В Гусев, Л.А. Минеева 1992, с изменениями)
|
Энергетический процесс |
Конечный акцептор электронов |
Продукты восстановления |
|
Нитратное дыхание и нитрификация |
||
|
Сульфатное и серное дыхание |
||
|
“Железное ” дыхание |
||
|
Карбонатное дыхание |
СН, ацетат |
|
|
Фумаратное дыхание |
Сукцинат |
Свойство организмов переносить электроны на нитраты, сульфаты и карбонаты обеспечивает в достаточной степени полное окисление органического или неорганического вещества без использования молекулярного кислорода и обуславливает возможность получения большого количества энергии, чем при брожении. При анаэробном дыхании выход энергии только на 10% ниже. Чем при аэробном. Организмы, для которых характерно анаэробное дыхание, имеют набор ферментов электронтранспортной цепи. Но цитохромоксилаза в них заменяется нитратредуктазой (при использовании в качестве акцептора электронов нитрата) или аденилсульфатредуктазой (при использовании сульфата) или другими ферментами.
Организмы, способные осуществлять анаэробное дыхание за счет нитратов, - факультативные анаэробы. Организмы, использующие сульфаты в анаэробном дыхании, относятся к анаэробам.
Вывод
Органические вещества из не органических зеленое растение образует только на свету. Эти вещества используются растением только для питания. Но растения не только питаются. Они дышат, как все живые существа. Дыхание происходит непрерывно днем о ночью. Дышат все органы растения. Растения дышат кислородом, а выделяют углекислый газ, как животные и человек.
Дыхание растений может происходить, как в темноте, так и на свету. Значит, на свету в растении протекают два противоположных процесса. Один процесс - фотосинтез, другой - дыхание. Во время фотосинтеза создаются органические вещества из неорганических и поглощается энергия солнечного света. Во время дыхания в растении расходуются органические вещества. А энергия, необходима для жизнедеятельности, освобождается. На свету в процессе фотосинтеза растения поглощают углекислый газ и выделяют кислород. Вместе с углекислым газом растения на свету поглощают из окружающего воздуха и кислород, необходимый растениям для дыхания, но в гораздо меньших количествах, чем выделяются при образовании сахара. Углекислого газа при фотосинтезе растения поглощают гораздо больше, чем выделяют его придыхании. Декоративные растения в комнате при хорошем освещении выделяют днем значительно больше кислорода, чем поглощают его в темноте ночью.
Дыхание во всех живых органов растения происходит непрерывно. Когда прекращается дыхание, растение, так же как и животное погибает.
Список литературы
1. Физиология и биохимия сельскохозяйственных растений Ф50/Н.Н. Третьяков, Е.И. Кошкин, Н.М. Макрушин и др.; под. ред. Н.Н. Третьякова. - М.; Колос, 2000 - 640 с.
2. Биология в экзаменационных вопросах и ответах Л44/ Лемеза Н.А., Камлюк Л.В.; 7-е изд. - М.: Айрис-пресс, 2003. - 512 с.
3. Ботаника: Учеб. Для 5-6 кл. сред. Шк.-19-е изд./Перераб. А.Н. Сладковым. - М.: Просвещение, 1987. - 256 с.
Растения живут благодаря процессу дыхания, но в отсутствие кислорода некоторое время они могут жить за счет анаэробного дыхания. Анаэробное дыхание растений
включается, когда необходимый растению кислород потребляется из органических соединений, главным образом из сахара, который обычно и является исходным материалом при нормальном дыхании.
Распределение сахара при анаэробном дыхании
При анаэробном дыхании сахар распадается по схеме: С 6 Н 12 О 6 → 2С 2 Н 5 ОН+2СО 2 + 48ккал Как видно, углерод сахара лишь частично окисляется до углекислого газа, а остальной углерод восстанавливается до этилового спирта, так как кислород извне не поступает, а превращение сахара происходит только за счет перераспределения кислорода, находящегося в его молекуле. Энергии в случае анаэробного дыхания выделяется всего 48 ккал, тогда как при полном окислении - 686 ккал, (подробнее: ). Такая разница объясняется тем, что в спирте остается большое количество потенциальной энергии, поскольку окисление идет не до конца.Анаэробные условия
Однако растения не могут жить долго в анаэробных условиях . Для того чтобы получить такое же количество энергии, которое оно имеет при дыхании, при анаэробном дыхании растение должно затратить очень большое количество запасного вещества. Поэтому в анаэробных условиях растения быстро погибают от истощения и, кроме того, от отравления спиртом , накапливающимся в тканях. Поэтому процесс анаэробного дыхания для высших растений - только временная замена кислородного дыхания . Анаэробное дыхание наблюдается у растений, длительное время находящихся при избытке влаги в почве, при образовании корки на поверхности почвы и хранении зерна в больших кучах.Анаэробное дыхание для микроорганизмов
Для многих низших растений (микроорганизмов ) анаэробное дыхание служит основным процессом добывания необходимой для жизни энергии и может поддерживать их жизнь неограниченное время. В этом случае анаэробное дыхание называется брожением . Микроорганизмы используют для брожения не собственные запасы питательных веществ, как это имеет место у , а питательные вещества из окружающей их среды. Анаэробное дыхание у растений сходно со спиртовым брожением . В анаэробных условиях под воздействием ряда ферментов образуются промежуточные продукты те же, что и при брожении, в частности пировиноградная кислота . В аэробных условиях пировиноградная кислота полностью окисляется до углекислоты и воды, а в анаэробных условиях при спиртовом брожении она распадается до С0 2 и спирта. На схеме показано взаимоотношение между нормальным дыханием - аэробным и анаэробным - спиртовым брожением.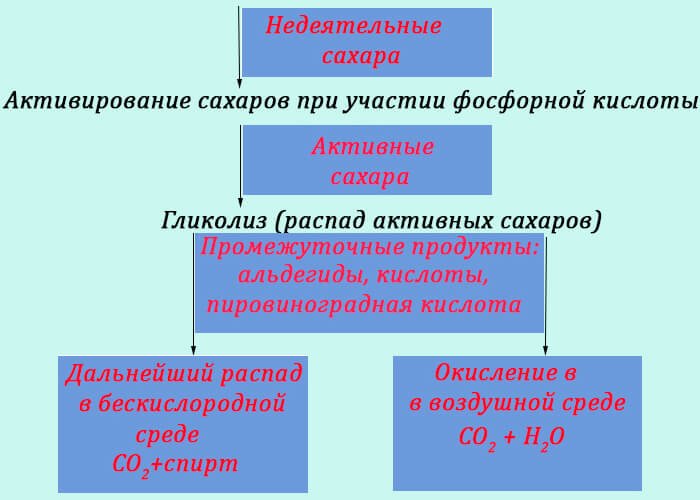 Аэробное и анаэробное дыхание.
Как видно из схемы, процессы дыхания и брожения одинаковы до образования пировиноградной кислоты. При дыхании для образования пировиноградной кислоты не требуется участия кислорода, т.е. эта фаза дыхания является анаэробной.
При доступе кислорода и наличии системы окислительных ферментов пировиноградная кислота окисляется до конца. При спиртовом брожении с участием фермента карбоксилазы карбоксил пировиноградной кислоты разрушается, выделяется углекислота и образуется уксусный альдегид
, которому при участии фермента дегидрогеназы передаются 2 атома водорода и он восстанавливается в этиловый спирт.
Таким образом, конечными продуктами спиртового брожения являются спирт и углекислота.
Аэробное и анаэробное дыхание.
Как видно из схемы, процессы дыхания и брожения одинаковы до образования пировиноградной кислоты. При дыхании для образования пировиноградной кислоты не требуется участия кислорода, т.е. эта фаза дыхания является анаэробной.
При доступе кислорода и наличии системы окислительных ферментов пировиноградная кислота окисляется до конца. При спиртовом брожении с участием фермента карбоксилазы карбоксил пировиноградной кислоты разрушается, выделяется углекислота и образуется уксусный альдегид
, которому при участии фермента дегидрогеназы передаются 2 атома водорода и он восстанавливается в этиловый спирт.
Таким образом, конечными продуктами спиртового брожения являются спирт и углекислота.
Последние материалы сайта
Жимолость

К чему снится старая обувь девушке
Сонник обувь. Любая обувь во сне означает дорогу, как в прямом, так и в переносном смысле. То есть, может указывать на символическую дорогу в отношениях с кем-либо, работе, деятельности. Например, снится новая обувь, если во сне она была красивой, добротн
Цветы

К чему снится мяч: толкование по разным сонникам
К чему снится мяч женщине: Походите по чиновничьим кабинетам. 1 Мяч по Соннику целительницы Федоровской Видеть мяч во сне значит: Если вам приснилось, что вы бросаете мячик - вскоре вас обидит близкий друг.Если вам приснилось, что вы вспарывает
Капуста

Ленивая лазанья с грибами Ленивая лазанья с лавашом
Ароматное, сытное итальянское блюдо – лазанью – можно сделать собственноручно дома. При этом можно замесить специальное тесто или поступить гораздо проще – заменить его лавашем. При правильных действиях лазанья из лаваша получается очень сочной, аппетитно
Томаты

Рис с мясом в духовке Рис запеченный в духовке с мясом
Рис с мясом в духовке, как и любое другое блюдо, имеет множество интерпретаций приготовления. В данном обзоре представлен самый простой вариант с минимальными трудозатратами. Пошаговый рецепт с фото. Видео-рецепт. Мясо с рисом - самое классическое со
Клубника
")
Как правильно выбрать паевой инвестиционный фонд (ПИФ)
г. Москва Компания "УПРАВЛЯЮЩАЯ ФИНАНСОВАЯ ИНВЕСТИЦИОННАЯ КОМПАНИЯ" зарегистрирована 29 мая 2014 года местным органом ФНС — Межрайонная инспекция Федеральной налоговой службы №46 по г. Москве. Полное официальное наименование — ОБЩЕСТВО С ОГРАНИЧЕННОЙ ОТ
Огурцы

Столбчатый фундамент своими руками — пошаговая инструкция
Практически все счастливые владельцы частных домов, коттеджей или дачных домиков хотели бы иметь на своем участке отдельно стоящую баню. Для ее возведения можно воспользоваться услугами профессиональных строителей или же соорудить баньку самостоятельно. О